






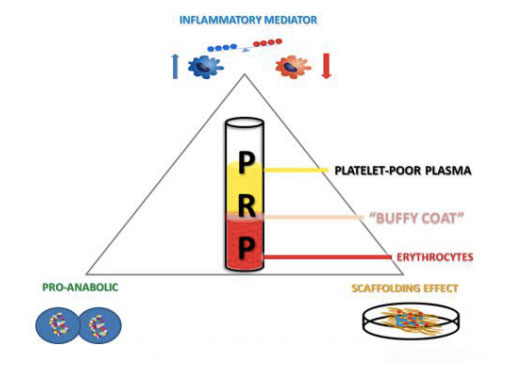
現在、PRP として知られる概念は 1970 年代に血液学の分野に初めて登場しました。血液学者は数十年前に、末梢血の基本値よりも高い血小板数から得られる血漿を表すために PRP という用語を作成しました。10 年以上後、PRP は多血小板フィブリン (PRF) の一種として顎顔面外科で使用されるようになりました。この PRP 誘導体中のフィブリンの含有量は、その接着性と定常状態の特性により重要な値を持ちますが、PRP は持続的な抗炎症特性を持ち、細胞増殖を刺激します。1990年代頃からようやくPRPが普及し始めました。最終的に、この技術は他の医療分野に移転されました。それ以来、この種のポジティブ生物学は広く研究され、プロスポーツ選手のさまざまな筋骨格系損傷の治療に応用され、メディアで広く注目されるようになりました。PRPは整形外科やスポーツ医学に効果があるほか、眼科、婦人科、泌尿器科、循環器科、小児科、形成外科でも使用されています。近年、PRP は皮膚潰瘍、傷跡修復、組織再生、皮膚の若返り、さらには脱毛の治療における可能性について皮膚科医からも賞賛されています。
PRP が治癒と炎症のプロセスを直接操作できるという事実を考慮すると、治癒カスケードを参考として導入する必要があります。治癒プロセスは次の 4 つの段階に分かれています。炎症;細胞とマトリックスの増殖、そして最終的には創傷のリモデリング。
組織の治癒
組織治癒カスケード反応が活性化され、血小板凝集、血栓の形成、および一時的な細胞外マトリックス (ECM) の発生が引き起こされます。次に、血小板が露出したコラーゲンと ECM タンパク質に付着し、α-顆粒に存在する生物活性分子の放出を引き起こします。血小板には、成長因子、化学療法因子、サイトカインなどのさまざまな生理活性分子のほか、プロスタグランジン、前立腺サイクリン、ヒスタミン、トロンボキサン、セロトニン、ブラジキニンなどの炎症誘発性メディエーターが含まれています。
治癒過程の最終段階は、創傷の再形成に依存します。組織のリモデリングは、同化反応と異化反応のバランスを確立するために厳密に規制されています。この段階では、血小板由来成長因子 (PDGF) およびトランスフォーミング成長因子 (TGF-β) フィブロネクチンとフィブロネクチンが線維芽細胞の増殖と遊走、および ECM 成分の合成を刺激します。ただし、創傷が成熟する時間は、創傷の重症度、個人の特徴、および損傷した組織の特定の治癒能力に大きく依存します。組織虚血、低酸素症、感染、成長因子の不均衡、さらにはメタボリックシンドローム関連疾患など、一部の病態生理学的要因や代謝要因が治癒過程に影響を与える可能性があります。
炎症誘発性微小環境は治癒過程を妨げます。さらに複雑なのは、高いプロテアーゼ活性が成長因子 (GF) の自然な作用を阻害することです。PRP は、有糸分裂、血管形成、走化性の特性に加えて、多くの成長因子の豊富な供給源でもあります。これらの生体分子は、炎症の増加を制御し、同化刺激を確立することにより、炎症組織における有害な影響に対抗する可能性があります。これらの特徴を考慮すると、研究者はさまざまな複雑な損傷の治療に大きな可能性を見出しているかもしれません。
多くの疾患、特に筋骨格系の疾患は、変形性関節症治療用の PRP など、炎症過程を調節する生物学的製剤に強く依存しています。この場合、関節軟骨の健康は、同化反応と異化反応の正確なバランスに依存します。この原則を念頭に置くと、特定のポジティブな生物学的薬剤の使用が健全なバランスを達成することに成功する可能性があります。PRP は血小板 α- を放出するため、顆粒に含まれる成長因子は、組織変化の可能性を調節するために広く使用されており、痛みも軽減します。実際、PRP 治療の主な目的の 1 つは、主要な炎症性および異化性微小環境を阻止し、抗炎症薬への変換を促進することです。他の著者は、トロンビン活性化 PRP がいくつかの生体分子の放出を増加させることを以前に実証しました。これらの因子には、肝細胞増殖因子(HGF)および腫瘍壊死因子(TNF-α)、トランスフォーミング増殖因子β1(TGF-β1)、血管内皮増殖因子(VEGF)および表皮増殖因子(EGF)が含まれます。他の研究では、PRP が II 型コラーゲンとアグリカン mRNA レベルの増加を促進し、同時にそれらに対する炎症誘発性サイトカイン インターロイキン – (IL) 1 の阻害を軽減することが示されています。HGF と TNF-α [28] のおかげで、PRP が抗炎症効果の確立に役立つ可能性があることも示唆されました。これらの分子製剤は両方とも、核因子κB (NF-κВ) の抗活性化活性と発現を減少させます。第二に、TGF-β 1 の発現によって単球の走化性も防止され、それによってケモカインのトランス活性化に対する TNF-α 効果が抑制されます。HGF は PRP によって誘発される抗炎症効果において不可欠な役割を果たしているようです。この強力な抗炎症性サイトカインは、NF-κ B シグナル伝達経路を破壊し、炎症誘発性サイトカインの発現によって炎症反応を阻害します。さらに、PRP は高レベルの一酸化窒素 (NO) を減らすこともできます。たとえば、関節軟骨では、NO 濃度の増加はコラーゲン合成を阻害し、軟骨細胞のアポトーシスを誘導する一方、マトリックスメタロプロテイナーゼ (MMP) の合成を増加させ、それによって異化作用の変換を促進することが証明されています。細胞変性の観点からは、PRP は特定の細胞型のオートファジーを操作できるとも考えられています。最終的な老化状態に達すると、一部の細胞グループは静的状態と自己再生の可能性を失います。しかし、最近の研究では、PRP 治療がこれらの有害な状態を十分に改善できることが示されています。Moussaらは、PRPがヒトの変形性関節症軟骨のアポトーシスを減少させながら、オートファジーと抗炎症マーカーを増加させることによって軟骨細胞の保護を誘導できることを証明した。ガルシア・プラットら。オートファジーが筋幹細胞の休止運命と老化運命の間の移行を決定することが報告されています。研究者らは、生体内では統合されたオートファジーが正常化することで細胞内損傷の蓄積が回避され、衛星細胞の老化や機能低下が防止されると考えている。最近のように、老化したヒト幹細胞においても、Parrish と Rodes は重要な貢献を果たし、PRP の抗炎症能力をさらに明らかにしました。今回は血小板と好中球の相互作用に焦点を当てます。研究者らは調査の中で、アラキドン酸によって放出された活性化血小板が好中球に吸収され、既知の炎症分子であるロイコトリエンとプロスタグランジンに変換されたと説明した。しかし、血小板好中球の相互作用により、ロイコトリエンはリポタンパク質に変換され、好中球の活性化を制限して透析を防ぎ、治癒カスケードの最終段階への遺伝を促進する効果的な抗炎症タンパク質であることが証明されています。
炎症誘発性微小環境は治癒過程を妨げます。さらに複雑なのは、高いプロテアーゼ活性が成長因子 (GF) の自然な作用を阻害することです。PRP は、有糸分裂、血管形成、走化性の特性に加えて、多くの成長因子の豊富な供給源でもあります。これらの生体分子は、炎症の増加を制御し、同化刺激を確立することにより、炎症組織における有害な影響を打ち消す可能性があります。
細胞因子
PRP のサイトカインは、組織修復のプロセスを操作し、炎症性損傷を制御する上で重要な役割を果たします。抗炎症性サイトカインは、主に活性化されたマクロファージによって誘導される炎症誘発性サイトカインの応答を媒介する広範囲の生化学分子です。抗炎症性サイトカインは、特定のサイトカイン阻害剤および可溶性サイトカイン受容体と相互作用して炎症を調節します。インターロイキン (IL) – 1 受容体拮抗薬である IL-4、IL-10、IL-11、および IL-13 は、主要な抗炎症薬であるサイトカインとして分類されます。さまざまな傷の種類に応じて、インターフェロン、白血病抑制因子、TGF-β、IL-6 などの一部のサイトカインは、炎症誘発性または抗炎症性の効果を示します。TNF-α、IL-1、IL-18 には特定のサイトカイン受容体があり、他のタンパク質の炎症促進効果を阻害する可能性があります [37]。IL-10 は最も効果的な抗炎症性サイトカインの 1 つであり、IL-1、IL-6、TNF-α などの炎症誘発性サイトカインを下方制御し、抗炎症因子を上方制御することができます。これらの抗調節機構は、炎症誘発性サイトカインの産生と機能において重要な役割を果たします。さらに、特定のサイトカインは、組織修復に重要な線維芽細胞を刺激する特定のシグナル応答を引き起こす可能性があります。炎症性サイトカイン TGF β 1、IL-1 β、IL-6、IL-13、および IL-33 は、線維芽細胞を刺激して筋線維芽細胞に分化し、ECM を改善します [38]。次に、線維芽細胞はサイトカイン TGF-β、IL-1 β、IL-33、CXC および CC ケモカインを分泌し、マクロファージなどの免疫細胞を活性化および動員することで炎症反応を促進します。これらの炎症細胞は、主に創傷除去の促進と、新しい組織の再構築に重要なケモカイン、代謝産物、成長因子の生合成の促進により、創傷において複数の役割を果たします。したがって、PRP のサイトカインは、細胞型媒介免疫応答を刺激し、炎症段階の退行を促進する上で重要な役割を果たします。実際、一部の研究者はこのプロセスを「再生炎症」と名付け、患者の不安にもかかわらず、炎症がシグナルを発するエピジェネティックなメカニズムを考慮すると、炎症段階は組織修復プロセスを正常に完了させるために必要かつ重要なステップであることを示しています。細胞の可塑性を促進します。
胎児の皮膚炎症におけるサイトカインの役割は、再生医療の研究にとって非常に重要です。胎児と成人の治癒メカニズムの違いは、胎児の年齢と関連する組織の種類に応じて、損傷した胎児の組織が時々元の状態に戻ることです。人間の場合、胎児の皮膚は 24 週間以内に完全に再生しますが、成人の場合、創傷治癒により瘢痕が形成されることがあります。私たちが知っているように、健康な組織と比較して、瘢痕組織の機械的特性は大幅に低下しており、その機能は制限されています。特にサイトカイン IL-10 に注目しています。IL-10 は羊水と胎児の皮膚で高度に発現していることがわかっており、サイトカインの多面発現効果によって促進され、胎児の皮膚の瘢痕のない修復に役割を果たすことが証明されています。ZgheibC et al.トランスジェニックノックアウト(KO)IL-10マウスおよび対照マウスへの胎児皮膚の移植が研究された。IL-10KO マウスは移植片周囲に炎症と瘢痕形成の兆候を示しましたが、対照群の移植片では生体力学特性に大きな変化は見られず、瘢痕治癒も見られませんでした。
抗炎症性サイトカインと炎症誘発性サイトカインの発現の間の微妙なバランスを調節することの重要性は、後者が過剰生産されると、最終的には特定の遺伝子の発現を低下させることによって細胞分解のシグナルを送ることになるということです。たとえば、筋骨格医学では、IL-1 β が軟骨の発達を担う SOX9 を下方制御します。SOX9 は、軟骨の発達に重要な転写因子を生成し、II 型コラーゲン アルファ 1 (Col2A1) を制御し、II 型コラーゲン遺伝子のコード化に関与しています。IL-1 β 最後に、Col2A1 とアグリカンの発現が減少しました。しかし、多血小板製品による治療は、IL-1 β を阻害することが示されています。コラーゲンをコードする遺伝子の発現を維持し、炎症誘発性サイトカインによって誘発される軟骨細胞のアポトーシスを軽減する再生医療の有力な味方です。
同化刺激:損傷した組織の炎症状態の調節に加えて、PRP のサイトカインは有糸分裂、化学的誘引、増殖の役割を果たすことで同化反応にも関与します。これは、Cavallo らによって主導された in vitro 研究です。ヒト軟骨細胞に対するさまざまな PRP の影響を研究する。研究者らは、血小板と白血球の濃度が比較的低い PRP 製品が正常な軟骨細胞の活動を刺激し、同化反応のいくつかの細胞機構の促進に役立つことを観察しました。たとえば、ii 型コラーゲンの発現とグリカンの凝集が観察されました。対照的に、高濃度の血小板と白血球は、さまざまなサイトカインが関与する他の細胞シグナル伝達経路を刺激すると考えられます。著者らは、これはこの特定の PRP 製剤中に多数の白血球が存在するためである可能性があると示唆しています。これらの細胞は、VEGF、FGF-b、インターロイキン IL-1b および IL-6 などの特定の成長因子の発現増加に関与していると考えられ、これらの因子が TIMP-1 および IL-10 を刺激する可能性があります。言い換えれば、「悪い」PRP 処方と比較して、血小板と白血球が豊富な PRP 混合物は軟骨細胞の相対的な浸潤を促進すると考えられます。
Schnabelらによって設計された研究。は、馬の腱組織における自家生体材料の役割を評価するために設計されました。著者らは、6頭の若い成馬(2~4歳)から血液と腱のサンプルを採取し、PRPを含む培地で培養した馬の浅指屈筋の腱外植片の遺伝子発現パターン、DNA、コラーゲン含有量の研究に焦点を当てた。または他の血液製剤。腱外植片を血液、血漿、PRP、血小板欠乏血漿 (PPP) または骨髄吸引液 (BMA) で培養し、100%、50%、または 10% の無血清 DMEM にアミノ酸を添加しました。…の後に適用可能な生化学分析を実行する際に、研究者らは、PRP 培地中の TGF-β PDGF-BB および PDGF-1 の濃度が、試験した他のすべての血液製剤の濃度よりも特に高かったことに注目しました。さらに、100% PRP 培地で培養した腱組織では、マトリックスタンパク質 COL1A1、COL3A1、および COMP の遺伝子発現の増加が示されましたが、異化酵素 MMPs3 および 13 は増加しませんでした。少なくとも腱の構造に関して、この in vivo 研究は、 autolo – 大型哺乳類の腱炎の治療用の痛風血液製剤 (PRP)。
チェンら。PRP の再建効果についてさらに議論しました。これまでの一連の研究で研究者らは、PRPが軟骨形成の促進に加えて、ECM合成の増加も促進し、関節軟骨と髄核の炎症反応を抑制することを証明した。PRP は Smad2/3-β シグナル経路のリン酸化を通じて TGF を活性化することができ、細胞の成長と分化において重要な役割を果たします。さらに、PRP 活性化後に形成されるフィブリン血餅は固体の三次元構造を提供し、細胞の接着を可能にし、新しい組織の構築につながる可能性があるとも考えられています。
他の研究者は、皮膚科学の分野で慢性皮膚潰瘍の治療に多大な貢献をしています。これも注目に値する。たとえば、2019年にヘスラー氏とシャム氏が実施した研究では、薬剤耐性の慢性潰瘍が依然として医療に多大な経済的負担をもたらしている一方で、PRPは実行可能で効果的な代替治療法として価値があることが示されている。特に、糖尿病性足部潰瘍はよく知られた重大な健康問題であり、四肢が切断されやすくなります。Ahmedらによって発表された研究。2017年の研究では、自家PRPゲルが必要な成長因子を放出することで慢性糖尿病足潰瘍患者の創傷治癒を促進し、それによって治癒率が大幅に向上する可能性があることを示した。同様に、Gonchar らは、糖尿病性足部潰瘍の治療改善における PRP と成長因子カクテルの再生可能性を検討し、議論しました。研究者らは、成長因子混合物の使用が解決策となる可能性が高く、これにより PRP と単一成長因子を使用する利点を改善できる可能性があると提案しました。したがって、単一の成長因子の使用と比較して、PRP と他の治療戦略の組み合わせは慢性潰瘍の治癒を大幅に促進する可能性があります。
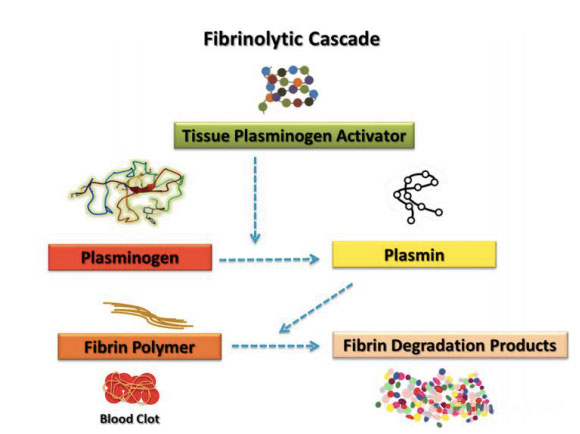
フィブリン
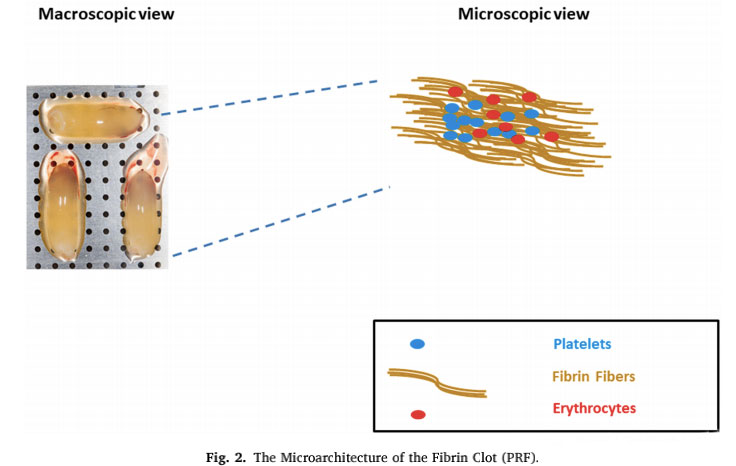
血小板は、線溶系に関連するいくつかの因子を担っており、線溶系反応をアップレギュレートまたはダウンレギュレートする可能性があります。血栓分解における血液成分と血小板機能の時間的関係と相対的な寄与は、依然としてコミュニティで広範に議論する価値のある問題です。文献には、治癒過程に影響を与える能力で有名な血小板のみに焦点を当てた多くの研究が紹介されています。数多くの優れた研究があるにもかかわらず、凝固因子や線溶系などの他の血液学的成分も効果的な創傷修復に大きく寄与することがわかっています。定義上、線維素溶解は、フィブリンの分解を促進する特定の酵素の活性化に依存する複雑な生物学的プロセスです。線維素溶解反応は、フィブリン分解産物 (fdp) が実際には組織修復の刺激に関与する分子因子である可能性があると他の著者によって提案されています。これまでの一連の重要な生物学的事象は、創傷治癒に必要なフィブリンの沈着と血管新生の除去から始まります。損傷後の血栓の形成は、組織を失血や微生物の侵入から保護する保護層として機能し、修復プロセス中に細胞が移動できる一時的なマトリックスとしても機能します。血栓はフィブリノーゲンがセリンプロテアーゼによって切断されることによって発生し、血小板は架橋されたフィブリン繊維メッシュに集まります。この反応は、血栓形成の主な出来事であるフィブリンモノマーの重合を引き起こしました。血餅は、活性化された血小板の脱顆粒中に放出されるサイトカインおよび成長因子の貯蔵庫としても使用できます。線溶系はプラスミンによって厳密に制御されており、細胞遊走、成長因子の生物学的利用能、および組織の炎症と再生に関与する他のプロテアーゼ系の制御の促進において重要な役割を果たしています。ウロキナーゼプラスミノーゲン活性化因子受容体(uPAR)やプラスミノーゲン活性化因子阻害剤-1(PAI-1)などの線維素溶解の主要な構成要素は、創傷治癒の成功に必要な特別な細胞型である間葉系幹細胞(MSC)で発現されることが知られています。 。
細胞の移動
uPA uPAR結合を介したプラスミノーゲンの活性化は、細胞外タンパク質分解を促進するため、炎症細胞の遊走を促進するプロセスです。膜貫通ドメインおよび細胞内ドメインが欠如しているため、uPAR は細胞遊走を制御するためにインテグリンやビテリンなどの共受容体を必要とします。さらに、uPA uPAR の結合により、ビトレクトネクチンおよびインテグリンに対する uPAR の親和性が増加し、細胞接着が促進されることが示されました。次に、プラスミノーゲン アクティベーター インヒビター 1 (PAI-1) が細胞を剥離させます。細胞表面のuPA uparインテグリン複合体のuPAに結合すると、uparビテリンとインテグリンビテリン間の相互作用を破壊します。
再生医療の文脈では、骨髄間葉系幹細胞は重篤な臓器損傷の場合に骨髄から動員されるため、多発骨折患者の循環中に見つかる可能性があります。ただし、末期腎不全、末期肝不全、心臓移植後の拒絶反応などの特定の場合には、これらの細胞が血液中に検出されない場合があります[66]。興味深いことに、これらのヒト骨髄由来の間葉(間質)前駆細胞は、健康な個人の血液中には検出されませんでした[67]。骨髄間葉系幹細胞 (BMSC) の動員における uPAR の役割は以前に提案されており、これは造血幹細胞 (HSC) の動員における uPAR の発生と同様です。ヴァラバネニら。その結果、uPAR欠損マウスにおける顆粒球コロニー刺激因子の使用がMSC不全を引き起こし、細胞遊走における線溶系の補助的な役割が再び強化されたことが示された。さらなる研究では、グリコシルホスファチジルイノシトールアンカー型 uPA 受容体が、次のような特定の細胞内シグナル伝達経路を活性化することによって接着、遊走、増殖、分化を調節していることも示されました: 生存可能なホスファチジルイノシトール 4,5-二リン酸 3-キナーゼ/Akt および ERK1/2 シグナル伝達経路、および接着キナーゼ(FAK)。
MSC 創傷治癒の文脈において、線維素溶解因子はそのさらなる重要性を証明しています。例えば、プラスミノーゲン欠損マウスは、創傷治癒事象に重大な遅延を示し、このプロセスにおいてプラスミンが重要であることを示した。人間の場合、プラスミンの損失は創傷治癒の合併症を引き起こす可能性もあります。血流の中断は組織の再生を大幅に阻害する可能性があり、糖尿病患者においてこれらの再生プロセスがより困難である理由もこのことが説明されています。
骨髄間葉系幹細胞が創傷部位に動員され、創傷治癒が促進されました。安定した条件下では、これらの細胞は uPAuPAR と PAI-1 を発現しました。最後の 2 つのタンパク質は、低酸素誘導因子 α (HIF-1 α) です。MSC 内の HIF-1 α は、FGF-2 および HGF の活性化によって FGF-2 および HGF のアップレギュレーションが促進されるため、ターゲティングは非常に便利です。HIF-2 α 次に、VEGF-A [77] が上方制御され、共に創傷治癒に貢献します。さらに、HGF は、相乗的に骨髄間葉系幹細胞の創傷部位への動員を促進すると思われます。虚血および低酸素状態は、創傷修復を著しく妨げることが示されていることに留意しなければならない。BMSC は低酸素レベルの組織に生息する傾向がありますが、損傷した組織で観察される悪条件下では移植細胞が死滅することが多いため、移植された BMSC の生体内での生存は制限されます。低酸素下での骨髄間葉系幹細胞の接着と生存の運命は、これらの細胞によって分泌される線維素溶解因子に依存します。PAI-1 はビテリンに対する親和性が高いため、uPAR およびインテグリンのビテリンへの結合を競合して、細胞の接着と遊走を阻害します。
単球と再生システム
文献によると、創傷治癒における単球の役割については多くの議論が行われています。マクロファージは主に血液単球に由来し、再生医療において重要な役割を果たしています[81]。好中球はIL-4、IL-1、IL-6、TNF-αを分泌するため、これらの細胞は通常、損傷後約24~48時間で傷口に浸透します。血小板はトロンビンと血小板因子 4 (PF4) を放出し、これにより単球の動員が促進され、マクロファージと樹状細胞に分化します。マクロファージの重要な特徴はその可塑性です。つまり、マクロファージは表現型を変換して内皮細胞などの他の細胞型に分化し、創傷微小環境におけるさまざまな生化学的刺激に対してさまざまな機能を示すことができます。炎症細胞は、刺激源としての局所分子シグナルに応じて、2 つの主要な表現型、M1 または M2 を発現します。M1 マクロファージは微生物因子によって誘導されるため、より炎症促進効果があります。対照的に、M2 マクロファージは通常 2 型反応によって生成され、抗炎症特性を持ち、通常は IL-4、IL-5、IL-9、および IL-13 の増加によって特徴付けられます。また、成長因子の生成を通じて組織の修復にも関与します。M1 サブタイプから M2 サブタイプへの移行は、主に創傷治癒の後期段階によって引き起こされます。M1 マクロファージは好中球のアポトーシスを引き起こし、これらの細胞のクリアランスを開始します)。好中球の食作用は、サイトカインの産生を停止させ、マクロファージを分極させ、TGF-β 1 を放出する一連の現象を活性化します。この成長因子は、筋線維芽細胞の分化と創傷収縮の重要な調節因子であり、炎症や炎症の解消を可能にします。治癒カスケードにおける増殖段階の開始[57]。細胞プロセスに関与するもう 1 つの高度に関連したタンパク質はセリン (SG) です。この造血細胞分泌顆粒プロテオグリカンは、マスト細胞、好中球、細胞傷害性 T リンパ球などの特定の免疫細胞に分泌タンパク質を貯蔵するのに必要であることがわかっています。多くの非造血細胞もプラスミノーゲンを合成しますが、すべての炎症細胞はこのタンパク質を大量に生成し、プロテアーゼ、サイトカイン、ケモカイン、成長因子などの他の炎症性メディエーターとさらに相互作用するために顆粒の中に蓄えます。SG に含まれる負に帯電したグリコサミノグリカン (GAG) 鎖は、細胞、タンパク質、および GAG 鎖に特異的な方法で本質的に帯電した顆粒成分に結合し、その貯蔵を促進できるため、分泌顆粒の安定性にとって重要であると考えられます。PRP 研究への参加に関して、Woulfe らは以前、SG 欠乏が血小板の形態変化と密接に関連していることを示した。血小板因子 4 β - トロンボグロブリンおよび血小板における PDGF 貯蔵の欠陥。in vitro では血小板の凝集と分泌が不十分であり、in vivo では血栓症が起こりません。したがって、研究者らは、このプロテオグリカンが血栓症の主な調節因子であると考えられると結論付けました。
多血小板製品は、収集と遠心分離を通じて個人の全血を取得し、その混合物を血漿、血小板、白血球、白血球を含むさまざまな層に分割することができます。血小板濃度が基本値よりも高い場合、副作用を最小限に抑えながら、骨と軟組織の成長を促進することができます。自己 PRP 製品の応用は比較的新しいバイオテクノロジーであり、さまざまな組織損傷の治癒を刺激し、促進するという楽観的な結果が継続的に示されています。この代替治療法の有効性は、生理学的創傷治癒および組織修復プロセスをシミュレートおよびサポートするための広範囲の成長因子およびタンパク質の局所送達に起因すると考えられます。さらに、線溶系は組織全体の修復に明らかに重要な影響を及ぼします。炎症細胞や骨髄間葉系幹細胞の細胞動員を変化させることに加えて、創傷治癒領域のタンパク質分解活性や、骨、軟骨、筋肉などの中胚葉組織の再生プロセスも調節できるため、重要な成分です。筋骨格系の医学。
治癒の促進は、医療分野の多くの専門家によって強く追求されている目標です。PRP はポジティブな生物学的ツールであり、再生イベントのカスケードを刺激し調整する上で有望な開発を提供し続けています。しかし、この治療ツールは依然として非常に複雑であるため、特に無数の生理活性因子とそれらのさまざまな相互作用メカニズムおよびシグナル伝達効果を放出するため、さらなる研究が必要です。
(本記事の内容は転載であり、当社は本記事に含まれる内容の正確性、信頼性、完全性について明示的、黙示的を問わずいかなる保証も行うものではなく、また本記事の見解については責任を負いません、ご了承ください。)
投稿日時: 2022 年 12 月 16 日